Here's a brief description (more detail can be found in Reeve & Webb 2002) Each neuron is modelled as an RC circuit with a fixed membrane capacitance and membrane conductance and a base potential across the membrane to which it will decay exponentially in the absence of external input. If the membrane potential rises above a threshold, the neuron will 'fire', sending a spike to any output synapses. The synapses are modelled as a variable conductance with a battery potential which corresponds to the reversal potential of the ion channel opened by the synaptic neurotransmitter. Synapses are characterised by a reversal potential, a delay (corresponding to the sum of possible axonal, neurotransmitter, and dendritic delays), a variable time course for the exponential decay of conductance, as well as a more standard 'weight' determining the standard conductance change in the affected ion channel. They also allow short-term adaptation of the weight. Thus this is a substantially more realistic model that standard ANN approaches, yet still runs in real time for robot control.
Using this system, and drawing on cricket neurobiology, we have designed
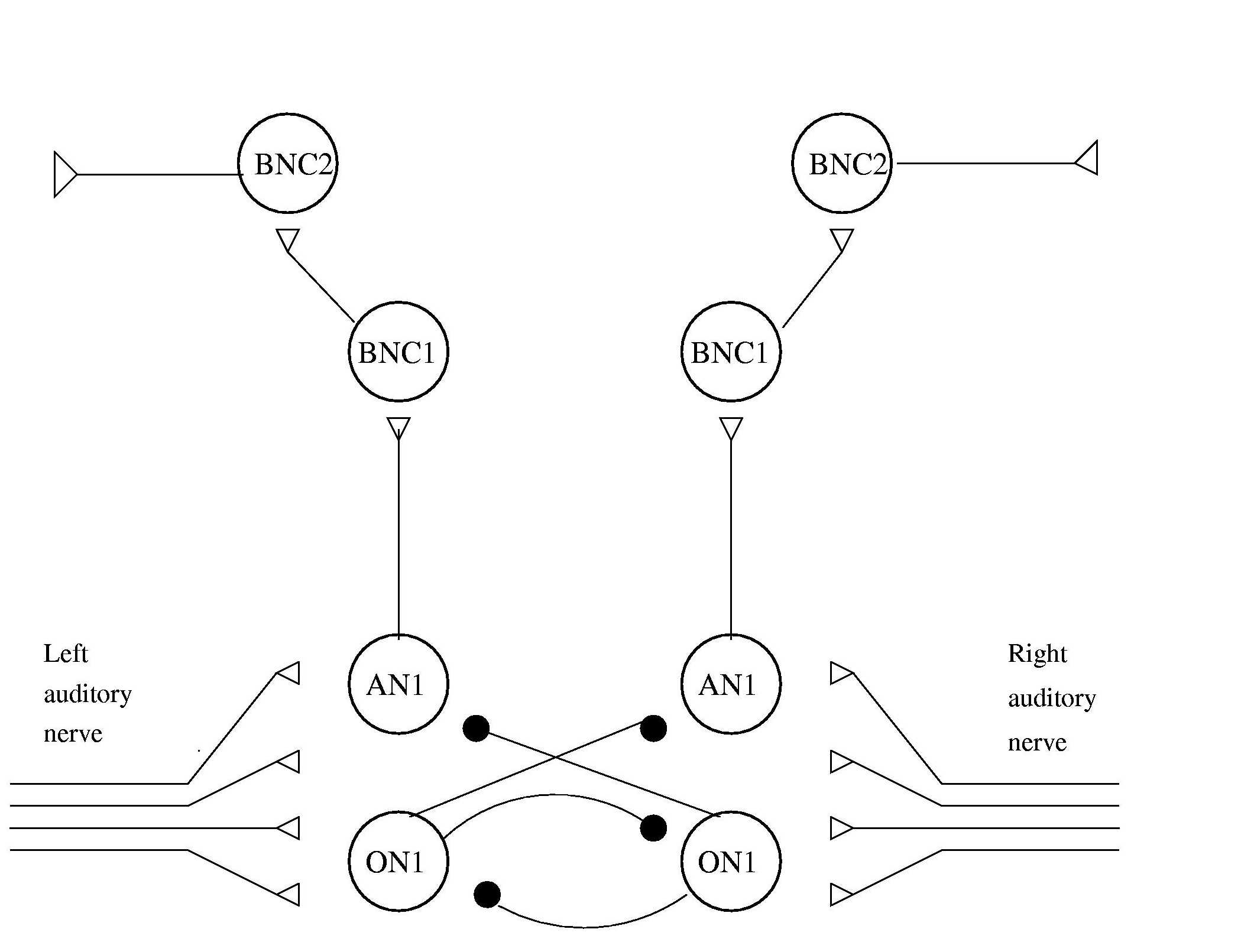
the following neural circuit to control the robot's behaviour:
The input to the left and right auditory interneurons consists of eight 'parallel fibres' from the ear sensors which encode the sound in poisson distributed spike trains proportional to the amplitude. A second pair of auditory interneurons receive the same input but provide cross-inhibition (based on identified neurons in the cricket, Horsemann & Huber, 1994) which serves to sharpen the difference and also acts as a gain control mechanism. The output from the auditory neurons passes through two levels of 'brain' neurons (based on those described by Schildberger, 1984) which filter for the temporal pattern. The output of these neurons is a spike, approximately once per chirp, on the side corresponding to the loudest and clearest sound pattern, indicating a turn in that direction is needed to approach the sound.
In previous work (Webb & Scutt, 2000) we used a simpler, but less realistic, neural model with only four neurons:
